
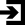

Доктор технических наук В. МАЙОРОВ.
Чуть более четверти века назад в журнале "Наука и жизнь" (№ 1, 1978 г.) была опубликована статья "Загадка запаха". Ее автор, кандидат химических наук Г. Шульпин, справедливо отмечал, что современное ему состояние науки о запахах примерно такое же, как состояние органической химии в 1835 году. Тогда один из зачинателей этой науки, Ф. Велер, писал, что органическая химия представляется ему дремучим лесом, из которого невозможно выбраться. Но уже через четверть века А. М. Бутлеров, создав теорию химического строения вещества, сумел "выбраться из чащи". Шульпин выражал уверенность, что загадка запаха будет решена едва ли не быстрее, чем в случае органической химии.
И он оказался прав на все 100%! В последнее время произошел настоящий прорыв в понимании молекулярных основ обоняния. Разберем основные стадии восприятия запахов в свете современных представлений.
КАК ВОСПРИНИМАЕТСЯ ЗАПАХ
Проделаем простой опыт. Возьмем флакон с пахучей жидкостью, например духами, откроем пробку и понюхаем содержимое в спокойном ритме дыхания. Легко обнаружить, что мы ощущаем запах только во время вдоха; начинается выдох - запах исчезает.
При вдохе через нос воздух вместе с молекулами пахучего вещества (называемого обонятельным стимулом или одорантом) проходит в каждой из двух носовых полостей по щелевидному каналу сложной конфигурации, который образован продольной носовой перегородкой и тремя носовыми раковинами. Здесь воздух очищается от пыли, увлажняется и нагревается. Затем часть воздуха поступает в расположенную в верхней задней зоне канала обонятельную область, имеющую вид щели, покрытой обонятельным эпителием.
Общая поверхность, занимаемая эпителием в обеих половинках носа взрослого человека, невелика - 2 - 4 см2 (у кролика эта величина равна 7-10 см2, у собак - 27 - 200 см2). Эпителий покрыт слоем обонятельной слизи и содержит три типа первичных клеток: обонятельные рецепторы, опорные и базальные клетки. Влекомые воздухом пахучие молекулы проникают в носовую полость и переносятся над поверхностью эпителия. При нормальном спокойном дыхании вблизи обонятельного эпителия проходит 7 -10% вдыхаемого воздуха. Обонятельный эпителий имеет толщину приблизительно 150-300 мкм. Он покрыт слоем слизи (10-50 мкм), который молекулам одоранта предстоит преодолеть, прежде чем они провзаимодействуют со специальными сенсорными нейронами - обонятельными рецепторами.
Основная функция обонятельного рецептора состоит в выделении, кодировании и передаче информации об интенсивности, качестве и продолжительности запаха в обонятельную луковицу и специальным центрам в головном мозге. Эпителий в обеих носовых полостях у человека содержит приблизительно 10 млн обонятельных нейронов ( у кролика - около 100 млн, а у немецкой овчарки - до 225 млн).
Как известно, нейрон состоит из тела и отростков: аксонов и дендритов. Нервный импульс с одной нервной клетки на другую передается с аксона на дендрит. Диаметр утолщенной центральной части обонятельного нейрона (сомы) 5-10 мкм. Дендритная часть в виде волокнистых отростков диаметром 1-2 мкм выходит к внешней поверхности эпителия. Здесь дендриты заканчиваются утолщением, от которого отходит пучок из 6-12 ресничек (цилий) диаметром 0,2-0,3 мкм и длиной до 200 мкм, погруженный внутрь слоя слизи (у кролика число ресничек в одном рецепторном нейроне составляет 30-60, а у собак достигает 100-150). Отходящее от сомы нервное волокно (аксон) имеет диаметр около 0,2 мкм и выходит к внутренней поверхности эпителия. Здесь аксоны от соседних нейронов объединяются в жгуты (филы), доходящие до обонятельной луковицы.
СЕМИОТИКА ОБОНЯНИЯ
Для того чтобы обонятельный сигнал был воспринят нейроном, молекула одоранта связывается со специальной белковой структурой, расположен ной в нейрональной клеточной мембране. Такая структура называется рецепторным белком. Используя методы молекулярной биологии, американские ученые Линда Бак и Ричард Аксель в 1991 году установили, что обонятельные нейроны у млекопитающих содержат около 1000 различных видов рецепторных белков (у человека их меньше - около 350). Признанием важности этого открытия стало присуждение им в 2004 году Нобелевской премии за исследования в области физиологии и медицины (см. "Наука и жизнь" № 12, 2004 г).
Каким образом рецепторы распределяются по нейронам: имеются ли отдельные представители этого семейства во всех обонятельных нейронах или каждый нейрон несет на своей мембране только один вид рецепторного белка? Как может мозг определить, какой из 1000 типов рецепторов подал сигнал? Имеющиеся данные позволяют сделать заключение о том, что на одном нейроне присутствует только обонятельный рецепторный белок одного вида. Нейроны с разными рецепторами обладают различной функциональностью, то есть в эпителии имеются тысячи различных типов нейронов. В этом случае проблема идентификации активированного запахом отдельного рецептора сводится к задаче выявления подавшего сигнал нейрона.
Принимая во внимание, что общее число обонятельных нейронов у человека около 10 млн, число обонятельных рецепторов одного типа исчисляется в среднем десятками тысяч.
Обонятельная система использует комбинаторную схему для идентификации одорантов и кодирования сигнала. Согласно ей один тип обонятельных рецепторов активируется множеством одорантов и один одорант активирует множество типов рецепторов. Различные одоранты кодируются различными комбинациями обонятельных рецепторов, причем увеличение концентрации стимула приводит к возрастанию числа активируемых рецепторов и к усложнению его рецепторного кода. В этой схеме каждый рецептор выступает в качестве одного из компонентов комбинаторного рецепторного кода для многих одорантов и как бы выполняет роль буквы своеобразного алфавита, из совокупности которых составляются соответствующие слова-запахи.
Минимальные структурные отличия молекул одорантов, например, по функциональной группе, по длине углеродной цепи, по пространственной структуре приводят к различному рецепторному коду. Для отличительного признака молекулы одоранта, способного изменить кодировку запаха, был предложен термин "одотоп" (odotope), или детерминант запаха. Различные обонятельные рецепторы, которые распознают один и тот же одорант, могут идентифицировать различные его признаки-одотопы. Одиночный обонятельный рецептор способен "различать" молекулы, отличающиеся длиной углеродной цепочки всего лишь на один атом углерода, или молекулы, имеющие одинаковую длину углеродной цепочки, но отличающиеся функциональной группой. Учитывая, что в эпителии млекопитающих имеется приблизительно 1000 видов обонятельных рецепторов, можно полагать, что такая комбинаторная схема позволяет различить громадное число одорантов (даже человек различает до 10 000 запахов).
Полученные в последнее время результаты экспериментальных исследований свойств обонятельных рецепторных белков позволили создать на молекулярном уровне структурную модель спиральной молекулы обонятельного белка. Обонятельные рецепторные белки принадлежат к суперсемейству мембранносвязанных рецепторов. Они пересекают двухслойную липидную мембрану реснички семь раз. У содержащей 300-350 аминокислот молекулы рецепторного белка три наружные петли соединяются с тремя внутриклеточными петлями семью пересекающими мембрану трансмембранными участками.
НЕОБХОДИМАЯ СЛИЗЬ
Находящиеся в потоке воздуха молекулы одоранта, перед тем как достичь обонятельных рецепторных нейронов, должны пересечь обволакива ющий поверхность обонятельного эпителия слой слизи. Физиологические функции слоя слизи полностью до сих пор не выяснены. Не вызывает сомнения, что она создает гидрофильную оболочку для чувствительных и хрупких обонятельных рецепторов, выполняя защитную функцию. Ведь систему восприятия сигнала нужно защитить от воздействия внешней среды, то есть от молекул одорантов, среди которых могут быть достаточно опасные и химически активные вещества.
Слой слизи состоит из двух подслоев. Внешний, водный, имеет толщину примерно 5 мкм, а внутренний, более вязкий, - около 30 мкм. Реснички-цилии направлены наклонно к внешней поверхности слоя слизи. Они образуют своего рода сетку с нерегулярными ячейками, причем эта сетка размещена у поверхности раздела подслоев так, что основная часть поверхности ресничек (около 85%) оказывается расположен ной вблизи границы раздела.
Слой слизи содержит разнообразные растворимые в воде белки, значительную часть которых составляют так называемые гликопротеины. Благодаря разветвленной молекулярной структуре эти белки способны связывать и удерживать молекулы воды, образуя гель.
Другие виды белков, содержащихся в слизи, взаимодействуют с молекулами одорантов и тем самым могут оказывать влияние на восприятие и распознавание запахов. Эти белки подразделяются на два основных класса - одорант-связующие белки (OBP) и одорант-разрушающие ферменты.
ОВР относятся к семейству белков, имеющих складчатую бочкообразную структуру с внутренней глубокой полостью, в которую попадают маленькие молекулы гидрофильных (жирорастворимых) одорантов. Разные подвиды этих белков отличаются высокой избирательностью взаимодействия с одорантами различных химических классов.
Полагают, что OBP способствуют растворению одоранта и транспортируют его молекулы сквозь слой слизи, действуют как фильтр для разделения одорантов, могут облегчать связывание одоранта с рецепторным белком и даже очищать околорецепторное пространство от ненужных компонентов.
Кроме одорант-связующих белков в слизи обонятельного эпителия вблизи рецепторных нейронов обнаружены несколько видов одорант-разрушающих ферментов. Все эти ферменты запускают реакции превращения молекул одорантов в другие соединения. Образующиеся в результате этих реакций продукты также вносят свой вклад в восприятие запаха. В конечном итоге все поступающие в слой слизи молекулы одорантов быстро, практически одновременно с завершением вдоха, теряют свою "запаховую" активность. Так что обонятельная система при каждом вдохе получает новую информацию от свежих порций одоранта.
ОБОНЯНИЕ НА УРОВНЕ МОЛЕКУЛ
Многие свойства системы восприятия запахов можно объяснить на молекулярном уровне. Молекула одоранта встречает на поверхности слизи, покрывающей обонятельный эпителий, молекулу одорант-связующего белка, которая связывает и переносит молекулу одоранта через слой слизи к поверхности реснички обонятельного нейрона. В ресничках осуществляется основной процесс передачи обонятельного сигнала. Его механизм достаточно типичен для многих видов взаимодействий физиологически активных веществ с рецепторами нервных клеток.
Молекула одоранта прикрепляется к определенному обонятельному рецептору (R). Между процессом связывания молекулы одоранта с рецептором и передачей обонятельного сигнала в нервную систему лежит сложный каскад биохимических реакций, проходящих в нейроне. Связывание молекулы одоранта с рецепторным белком активирует так называемый G-белок, расположенный на внутренней стороне клеточной мембраны. G-белок в свою очередь активирует аденилатциклазу (AC) - фермент, преобразующий внутриклеточный аденозинтрифосфат (ATP) в циклический аденозинмонофосфат (cAMP). А уже cAMP активирует другой мембранносвязанный белок, который называется ионным каналом, поскольку открывает и закрывает вход заряженным частицам внутрь клетки. Когда ионный канал открыт, в клетку проникают катионы металлов. Таким способом меняется электрический потенциал клеточной мембраны и генерируется электрический импульс, передающий сигнал с одного нейрона на другой.
Несколько молекулярных стадий передачи внутриклеточного сигнала обеспечивают его усиление, в результате чего небольшого числа молекул одоранта становится достаточно для генерирования нейроном электрического импульса. Такие усилительные каскады обеспечивают большую чувствительность системы восприятия запахов.
Итак, активация рецепторного белка молекулой одоранта в конечном счете приводит к генерированию электрического тока в обонятельном рецепторном нейроне. Ток распространяется по дендриту нейрона в его соматическую часть, где возбуждает выходной электрический импульс. Этот импульс передается по нейрональному аксону в обонятельную луковицу.
Одиночный электрический сигнал-импульс на выходе имеет длительность не более 5 мс и пиковую амплитуду около 100 мкВ. Почти все нейроны генерируют импульсы и при отсутствии воздействия одоранта, то есть обладают спонтанной активностью, называемой биологическим шумом. Частота этих импульсов меняется в диапазоне от 0,07 до 1,8 импульса в секунду.
ЛУКОВИЧНАЯ НЕЙРОСЕТЬ
Обонятельные рецепторные нейроны распознают громадное число разнообразных молекул пахучих веществ и посылают информацию о них через аксоны в обонятельную луковицу, служащую первым центром обработки обонятельной информации в головном мозге. Парные обонятельные луковицы представляют собой продолговатые образования "на ножках". Отсюда начинается путь обонятельного сигнала к полушариям мозга. Аксоны обонятельных нейронов оканчиваются в обонятельной луковице разветвлениями в сферических концентраторах (диаметром 100-200 мкм), называемых гломерулами. В гломерулах осуществляется контакт между окончаниями аксонов обонятельных нейронов и дендритами нейронов второго порядка, которыми являются митральные и пучковые клетки.
Митральные клетки - самые крупные нервные клетки, выходящие из обонятельной луковицы. Пучковые клетки меньше митральных, но функционально с ними схожи. Представление о количестве нервных клеток у млекопитающих могут дать характеристики обонятельной системы кролика. В ней имеется по 50 миллионов обонятельных рецепторных нейронов справа и слева (ровно в десять раз больше, чем у человека). Аксоны обонятельных рецепторов распределены между 1900 гломерулами обонятельной луковицы - примерно по 26 000 аксонов на гломерулу. Дендритные окончания 45 000 митральных и 130 000 пучковых клеток получают сигналы от аксонов в гломерулах и передают их из обонятельной луковицы в центры обоняния в головном мозге. Около 24 митральных и 70 пучковых клеток получают информацию от аксонов в каждой гломеруле. У человека около 10 млн аксонов обонятельных нейронов распределяются по 2000 гломерул обонятельной луковицы.
Все аксоны одной популяции обонятельных нейронов сходятся на две гломерулы, зеркально расположенные по разные стороны двумерного поверхностного слоя обонятельной луковицы. В зависимости от содержания передаваемого сигнала гломерулы активируются различным образом. Совокупность активированных гломерул называется картой запаха и представляет своего рода "слепок" запаха, то есть она показывает, из каких пахучих веществ состоит воспринимаемый обонятельный объект.
Механизм активации гломерул до сих пор не выяснен. Усилия исследователей направлены на то, чтобы выяснить, каким образом многообразие одорантов воспроизводится в двумерном слое гломерул на поверхности обонятельной луковицы. Кстати, эти отображения имеют динамический характер - они постоянно меняются в ходе восприятия запаха, усложняя научную задачу.
Обонятельная луковица - это большая многослойная нейросеть для пространственно-временнoй обработки отображения запаха в гломерулах. Ее можно рассматривать как совокупность множества микросхем с большим количеством связей, со взаимной активацией и ингибированием активности нейронов. Выполняемые нейронами операции выделяют характерные свойства карты запаха.
От обонятельной луковицы аксоны митральных и пучковых клеток передают информацию в первичные обонятельные участки коры головного мозга, а затем в высшие ее участки, где формируется осознанное ощущение запаха, и в лимбическую систему, которая порождает эмоциональную и мотивационную реакцию на обонятельный сигнал.
Свойства обонятельных зон коры головного мозга позволяют формировать ассоциативную память, которая устанавливает связь нового аромата с отпечатками воспринятых ранее обонятельных стимулов. Полагают, что процесс идентификации одоранта включает сравнение получающегося отображения с его описанием в семантической памяти. В случае совпадения отпечатка и памяти о запахе происходит какой-либо ответ (эмоциональный, двигательный) организма. Процесс этот осуществляется очень быстро, в течение секунды, и информация о совпадении после ответа сразу сбрасывается, поскольку мозг готовит себя к решению следующей задачи восприятия запаха.
ЗАГАДКИ ЗАПАХОВ
То, о чем говорилось в предыдущих разделах, относится пусть к самому сложному, основополагающему, но начальному разделу науки о запахах - к их восприятию. Не раскрыт механизм взаимодействия обоняния с другими системами восприятия, например со вкусом (см. "Наука и жизнь" № 8, 2003 г., с. 16-20). Ведь известно, что если человеку зажать ноздри, то при дегустации даже хорошо известных вкусовых пищевых продуктов (например - кофе) он не в состоянии точно определить, что он пробовал. Достаточно разжать ноздри - и вкусовые ощущения восстанавливаются.
С молекулярной точки зрения пока непонятно, в каких единицах измерять интенсивность запаха и от чего она зависит, что такое качество запаха, его "букет", чем отличается один запах от другого и как охарактеризовать это отличие, что происходит с запахом при смешивании различных одорантов. Оказывается, что независимо от вида одорантов и уровня подготовленности даже опытный эксперт не может определить все составляющие смесь компоненты, если их больше трех. Если же смесь содержит более десяти одорантов, то человек не в состоянии идентифицировать ни одного из них.
Остается еще множество вопросов, касающихся механизмов и видов воздействия запахов на эмоциональное, психическое и физическое состояния человека. В последнее время на эту тему появилось немало спекуляций, чему поспособствовал вышедший в 1985 году роман П. Зюскинда "Парфюмер", более восьми лет прочно занимавший место в первой десятке бестселлеров на западном книжном рынке. Фантазии на тему чрезвычайной силы подсознательного воздействия ароматов на эмоциональное состояние человека обеспечили этому произведению огромный успех.
Однако художественный вымысел постепенно получает обоснование. Недавно в периодической печати появились сообщения о том, что американские военные "парфюмеры" разработали на редкость дурно пахнущую бомбу, способную не только вызвать отвращение, но и разогнать солдат противника или агрессивно настроенную толпу.
Общественные аллюзии на парфюмерные темы подстегнули всеобщий интерес к искусству ароматерапии. Расширилось использование ароматов в общественных местах, таких, как офисы, торговые залы, холлы гостиниц. Появились даже специальным образом ароматизированные товары, улучшающие настроение. Возникла такая отрасль рыночной экономики, как аромамаркетинг - "наука" о привлечении клиентов с помощью приятных запахов. Так, запах кожи навевает покупателю мысли о дорогом качественном товаре, аромат кофе побуждает к покупкам для домашнего ужина и т.д. Каким образом запахи формируют в головном мозге сигналы, побуждающие человека совершать покупки? Ученым предстоит совершить еще немало открытий, прежде чем ответить на этот и многие другие вопросы и отделить мифы о запахах от реальности.
Литература
Лозовская Е., канд. физ.-мат. наук. Штрих-код запаха // Наука и жизнь, 2004, № 12.
Майоров В. А. Запахи: их восприятие, воздействие, устранение. - М.: Мир, 2006.
Марголина А., канд. биол. наук. Сладкая власть феромонов // Наука и жизнь, 2005, № 7.
Шульпин Г., канд. хим. наук. Загадка запаха // Наука и жизнь, 1978, № 1.
Пути передачи информации о запахах в головной мозг.
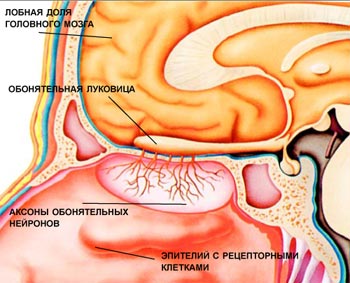
Схематическое изображение обонятельного эпителия. Базальные клетки являются клетками-предшественниками обонятельных рецепторных нейронов.
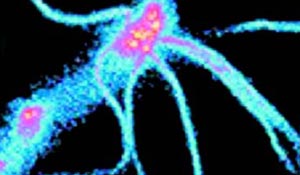
Изображение реснички обонятельного нейрона, сделанное с помощью флуоресцентного красителя.
На мембране ресничек расположены рецепторные белки, взаимодействующие с молекулами одорантов.
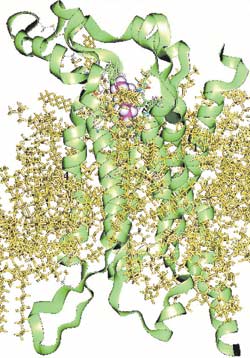
Модель молекулы обонятельного рецепторного белка мыши, к которому присоединена молекула одоранта - гексанола (пурпурного цвета).
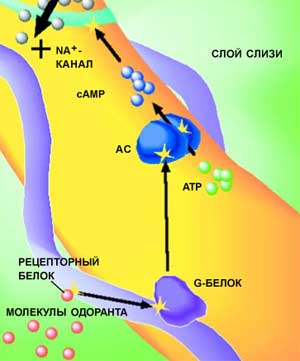
Одна из моделей процесса преобразования сигнала внутри реснички обонятельного нейрона.
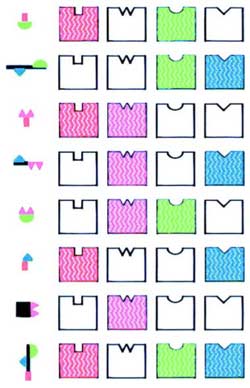
Схематическое изображение комбинаторных рецепторных кодов одорантов.
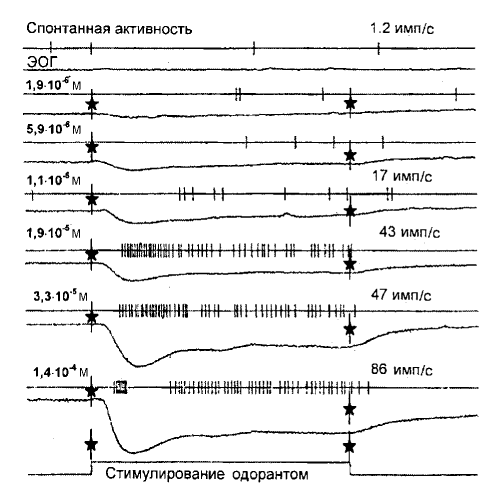
Электроольфактограмма (ЭОГ) - электрический колебательный сигнал, регистрируемый специальным электродом с участка внешней поверхности обонятельного эпителия крысы.
ПОДРОБНОЕ ОПИСАНИЕ ИЛЛЮСТРАЦИЙ
Пути передачи информации о запахах в головной мозг. В обонятельном эпителии в носовой полости расположены обонятельные рецепторные нейроны. Нервные волокна (аксоны) идут от нейронов на обонятельные луковицы, которые принимают и обрабатывают информацию о запахе. В луковице обонятельные нейроны передают сигнал на митральные клетки. Аксоны митральных клеток переносят сигналы о запахах в обонятельный центр мозга.
Пути передачи информации о запахах в головной мозг. В обонятельном эпителии в носовой полости расположены обонятельные рецепторные нейроны. Нервные волокна (аксоны) идут от нейронов на обонятельные луковицы, которые принимают и обрабатывают информацию о запахе. В луковице обонятельные нейроны передают сигнал на митральные клетки. Аксоны митральных клеток переносят сигналы о запахах в обонятельный центр мозга.
Одна из моделей процесса преобразования сигнала внутри реснички обонятельного нейрона. Молекула одоранта присоединяется к рецепторному белку. Связывание молекулы одоранта с рецепторным белком активирует так называемый G-белок, расположенный на внутренней стороне клеточной мембраны. G-белок активирует фермент аденилатциклазу (AC), который преобразует внутриклеточный аденозинтрифосфат (ATP) в циклический аденозинмонофосфат (cAMP). А cAMP, в свою очередь, активирует ионный канал, через который в клетку проникают катионы Na<SUP>+</SUP>. Таким способом в мембране генерируется электрический импульс, передающий сигнал с одного нейрона на другой.
Схематическое изображение комбинаторных рецепторных кодов одорантов. Молекулы различных одорантов (слева) распознаются несколькими рецепторами (справа). Молекулы имеют различные структурные свойства (одотопы, или детерминанты, обозначенные символами различного цвета и формы). В зависимости от детерминанты одорантам соответствуют различные комбинации рецепторов. Каждый рецептор выполняет роль одного из компонентов комбинаторного рецепторного кода для многих одорантов.
Электроольфактограмма (ЭОГ) - электрический колебательный сигнал, регистрируемый специальным электродом с участка внешней поверхности обонятельного эпителия крысы. Постепенное повышение концентрации одоранта позволяет проследить динамику реакции нейронов. Обонятельный нейрон проявил умеренную спонтанную активность при отсутствии стимула - около 1,2 имп/с (верхняя пара записей). При наименьшей концентрации стимула (1,9x10<SUP>-6 </SUP>М) на ЭОГ появляется сигнал, но активность одиночного нейрона не возрастает. Повышение концентрации одоранта до 5,9x10<SUP>-6</SUP> М вызывает увеличение амплитуды сигнала на ЭОГ. При этом на выходе нейрона фиксируются четыре последовательных импульса. Это дает основание полагать, что пороговая концентрация обонятельного стимула для данного нейрона находится между 1,9x10<SUP>-6 </SUP>и 5,9x10<SUP>-6</SUP> М. Дальнейшее повышение концентрации одоранта увеличивает активность
Схематическое изображение комбинаторных рецепторных кодов одорантов. Молекулы различных одорантов (слева) распознаются несколькими рецепторами (справа). Молекулы имеют различные структурные свойства (одотопы, или детерминанты, обозначенные символами различного цвета и формы). В зависимости от детерминанты одорантам соответствуют различные комбинации рецепторов. Каждый рецептор выполняет роль одного из компонентов комбинаторного рецепторного кода для многих одорантов.
Электроольфактограмма (ЭОГ) - электрический колебательный сигнал, регистрируемый специальным электродом с участка внешней поверхности обонятельного эпителия крысы. Постепенное повышение концентрации одоранта позволяет проследить динамику реакции нейронов. Обонятельный нейрон проявил умеренную спонтанную активность при отсутствии стимула - около 1,2 имп/с (верхняя пара записей). При наименьшей концентрации стимула (1,9x10<SUP>-6 </SUP>М) на ЭОГ появляется сигнал, но активность одиночного нейрона не возрастает. Повышение концентрации одоранта до 5,9x10<SUP>-6</SUP> М вызывает увеличение амплитуды сигнала на ЭОГ. При этом на выходе нейрона фиксируются четыре последовательных импульса. Это дает основание полагать, что пороговая концентрация обонятельного стимула для данного нейрона находится между 1,9x10<SUP>-6 </SUP>и 5,9x10<SUP>-6</SUP> М. Дальнейшее повышение концентрации одоранта увеличивает активность

